Эпистаз
Эпистаз, с другой стороны, описывает влияние одного гена на другой. Термин “эпистаз” означает, что один ген маскирует или модифицирует проявление фенотипа другого гена. Таким образом, эпистатический ген подавляет экспрессию другого гена или изменяет его проявление, влияя на фенотип организма.
Например, у кошек гены, определяющие наличие или отсутствие пигмента меланина в шерсти, могут быть подавлены другим геном, контролирующим осветление шерсти. В этом случае эпистатический ген определяет окончательный цвет шерсти, не зависимо от генов, ответственных за ее изначальный цвет.
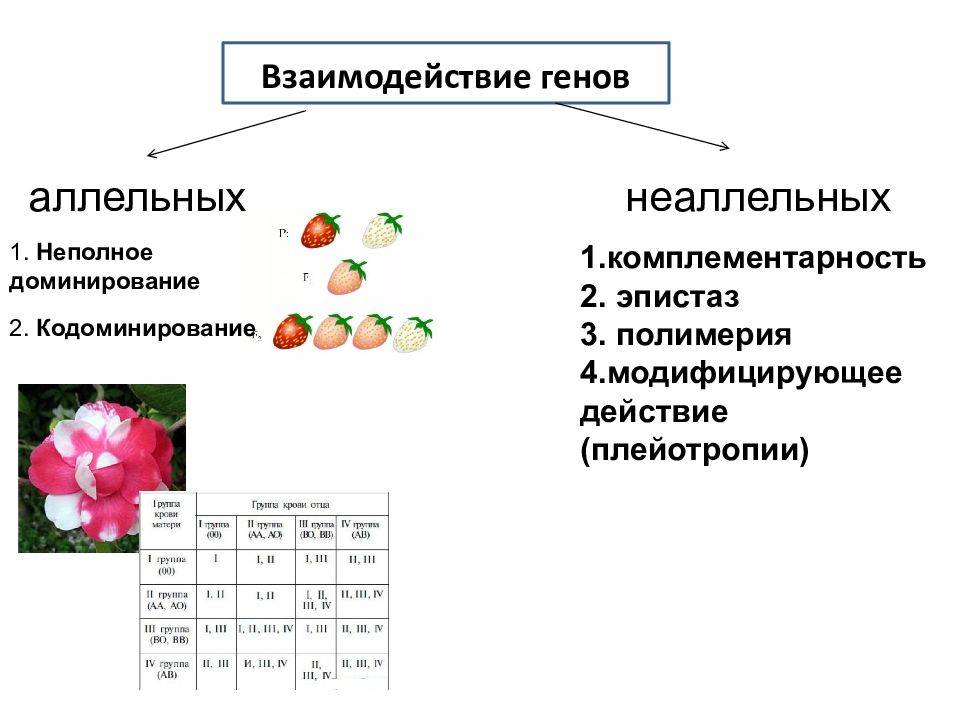
Типы взаимодействия аллельных генов
Различают три типа взаимодействия аллельных генов: полное доминирование, неполное доминирование, кодоминирование.
- Полное доминирование — явление, когда доминантный ген полностью подавляет работу рецессивного гена, в результате чего развивается доминантный признак.
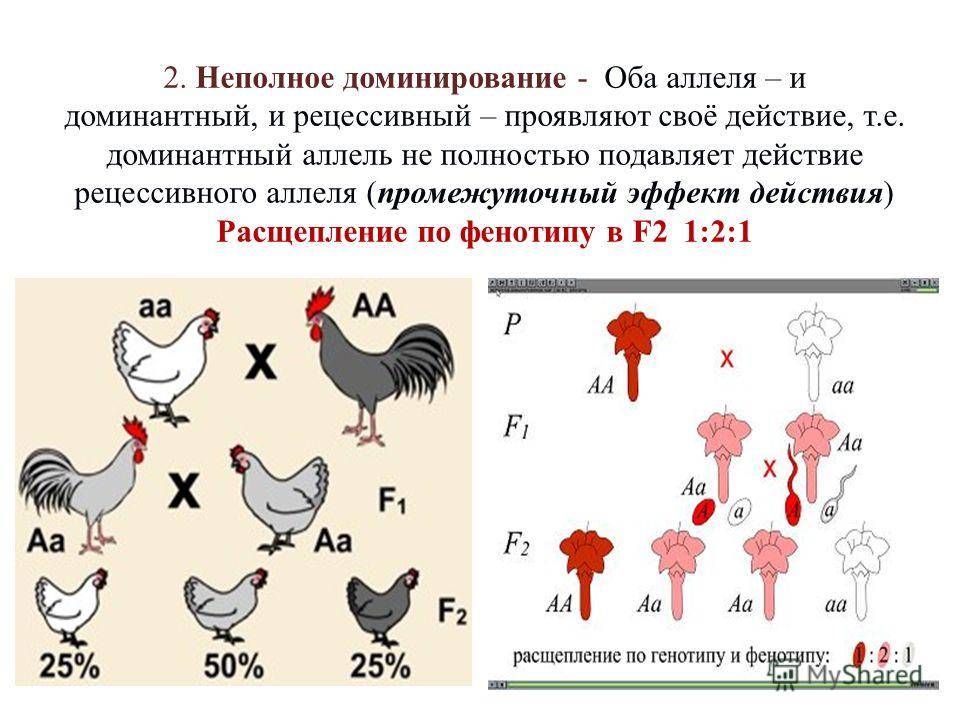
- Неполное доминирование — явление, когда доминантный ген не полностью подавляет работу рецессивного гена, в результате чего развивается промежуточный признак.
- Кодоминирование (независимое проявление) — явление, когда в формировании признака у гетерозиготного организма участвуют оба аллеля.
У человека с серией множественных аллелей представлен ген, определяющий группу крови. При этом гены, обусловливающие группы крови A и B, являются кодоминантными по отношению друг к другу и оба доминантны по отношению к гену, определяющему группу крови 0.
Что такое доминирование
Доминирование относится к феномену генетики, при котором у индивида, содержащего две аллельные формы определенного гена, одна выражается за исключением других. Как правило, ген, который определяет конкретный признак, существует в нескольких формах, известных как аллели. Аллели определенного гена расположены в одном и том же локусе гомологичных хромосом. Следовательно, диплоидный организм состоит из двух аллелей. Каждый аллель наследуется от каждого родителя. Каждый аллель также передается потомству половым путем. Наследование генов через поколения было впервые описано Грегором Менделем в 1890-х годах.
Парные аллели могут быть гетерозиготными или гомозиготными. Гомозиготные пары аллелей состоят из идентичных аллелей, тогда как гетерозиготные пары аллелей состоят из разных аллелей. Гомозиготные аллели выражают свой фенотип таким, какой он есть. Но в гетерозиготной паре аллелей один аллель экспрессируется над другим. Этот аллель известен как доминантный аллель. Присутствие доминантного аллеля в паре аллелей идентифицируется как доминантность. Можно выделить три типа моделей доминирования: полное доминирование, неполное доминирование и совместное доминирование.
Полное доминирование
При полном доминировании один аллель полностью доминирует над другим. Полностью маскированный аллель называется рецессивным аллелем. Таким образом, фенотип гена полностью определяется доминантным аллелем. Квадрат Пуннетта, описывающий менделевское наследование цвета цветка растения гороха, показан на Рисунок 1.

Рисунок 1: Наследование цвета растений гороха
Три генотипа, BB, Bb и bb, определяют цвет цветка у растений гороха. Доминирующим фенотипом для цветка является фиолетовый; белый рецессивный фенотип. Таким образом, доминантный аллель идентифицирован как B, тогда как рецессивный аллель идентифицирован как b.
Неполное Доминирование
При неполном доминировании один аллель не полностью доминирует над другим. Таким образом, ни доминантный, ни рецессивный фенотип не выражен. Но третий фенотип, который содержит смесь как доминантного, так и рецессивного фенотипа, выражен. Розовый цвет цветка львиного зева – пример неполного доминирования. Два фенотипа – красный и белый в цветах львиного зева.
Co-доминирование
При совместном доминировании ни один из аллелей не является доминантным. Однако оба аллеля полностью экспрессированы. Таким образом, в феномене доминантности экспрессируется более одного фенотипа. Наследование группы крови АВО у людей является примером совместного доминирования.
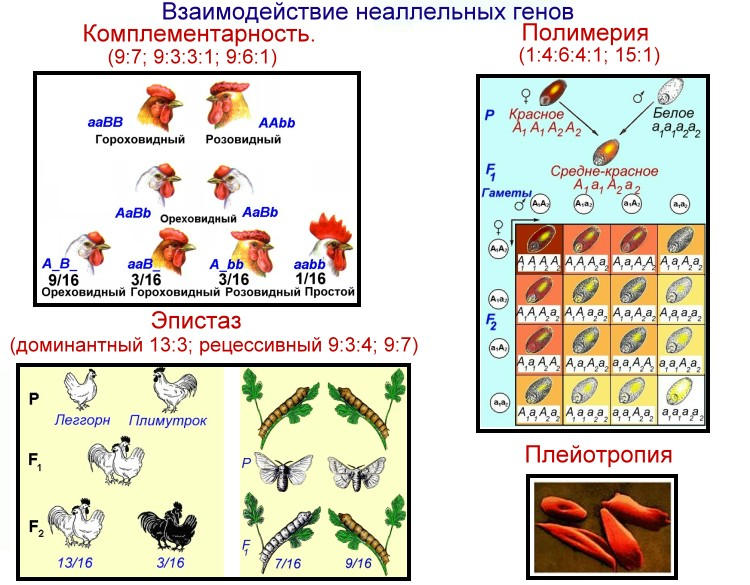
Взаимодействия неаллельных генов: комплементарность, эпистаз, полимерия
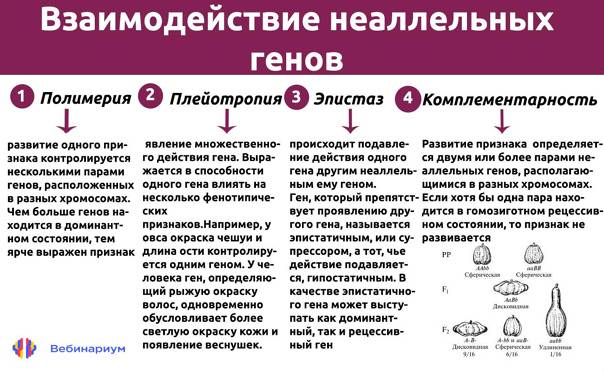
1. Комплементарность. Этот вид взаимодействия генов заключается в том, что при наличии двух доминантных аллелей разных генов появляется новый признак, то есть для появления нового признака у организма должен быть генотип АВ.
2. Эпистаз. При эпистатическом взаимодействии одна пара генов может подавлять действие другой пары генов.
3. Полимерия. Многие признаки определяются несколькими парами генов.
Это характерно, в основном, для количественных признаков, таких как яйценоскость у кур, жирность молока у коров.
4. Плейотропное действие гена. При плейотропном действии гена один ген определяет развитие или влияет на проявление нескольких признаков.
ГОМОЗИГОТНОСТЬ— состояние организма, при к-ром гомологичные хромосомы несут одну и ту же аллель того или иного гена.
Самооплодотворяющиеся организмы практитически гомозиготны.
Гетерозиготность — состояние, при котором гомологичные хромосомы несут разные формы (аллели) того или иного гена.
Гемизиготность —состояние, связанное с тем, что у организма один или несколько генов не парные, т. е. не имеют аллельных партнеров.
Закономерности наследования при моногибридном скрещивании.
Моногибридное скрещивание — скрещивание форм, отличающихся друг от друга по одной паре альтернативных признаков.
Первый закон Менделя (единообразия гибридов) – при скрещивании гомозиготных родительских особей, которые отличаются по одной паре аллельных признаков, все гибриды первого поколения единообразны по фенотипу и генотипу.
Второй закон Менделя (расщепления гибридов второго поколения) – при моногибридном скрещивании гетерозиготных организмов у гибридов второго поколения происходит расщепление по фенотипу в отношении 3:1 и по генотипу – 1:2:1
Дигибридное и полигибридное скрещивание.
Дигибридное скрещивание — скрещивание организмов, различающихся по двум парам альтернативных признаков.
Третий закон Менделя (независимого наследования признаков) – при скрещивании двух гомозиготных особей, отличающихся друг от друга по двум и более парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях.
Закон проявляется, как правило, для тех пар признаков, гены которых находятся в не гомологичных хромосомах.
Если обозначить буквой и число аллельных пар в негомологичных хромосомах, то число фенотипических классов будет определяться формулой 2n, а число генотипических классов — 3n. При неполном доминировании количество фенотипических и генотипических классов совпадает.
Цитологические основы законов Менделя базируются на:
1) парности хромосом (парности генов, обусловливающих возможность развития какого-либо признака)
2) особенностях мейоза (процессах, происходящих в мейозе, которые обеспечивают независимое расхождение хромосом с находящимися на них генами к разным полюсам клетки, а затем и в разные гаметы)
3) особенностях процесса оплодотворения (случайного комбинирования хромосом, несущих по одному гену из каждой аллельной пары)
Формула расщепления по обоим признакам. 9:3:3:1
Дигибридное и полигибридное скрещивание. Закон независимого комбинирования генов и его цитологические основы.
Общая формула расщепления при независимом наследовании.
Дигибридное скрещивание — скрещивание двух особей, отличающихся друг от друга по двум парам альтернативных признаков.
Полигибридное скрещивание — скрещивание двух особей, отличающихся друг от друга по двум и более парам альтернативных признаков.
Закон независимого комбинирования генов. Третий закон Менделя (независимого наследования признаков) – при скрещивании двух гомозиготных особей, отличающихся друг от друга по двум и более парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях.






Закон проявляется, как правило, для тех пар признаков, гены которых находятся в негомологичных хромосомах.
Цитологические основы:случайное комбинирование хромосом при оплодотворении, несущих по одному гену из каждой аллельной пары.
Общая формула расщепления при независимом наследовании.
По фенотипу: 9 (А-В-):3(А-вв):3(ааВ-):1(аавв).
Множественные аллели. Наследование групп крови человека системы AB0.
Множественные аллели – различные состояния (три или более) одного и того же локуса хромосомы, возникшие в результате мутаций.
Множественный аллелизм — наличие у гена множественных аллелей. Примером множественного аллелизма может служит система групп крови АВО.
Наследование групп крови. В локусе гена АВО возможны три варианта (аллеля) — 0, A и B, которые экспрессируются по аутосомно-кодоминантному типу.
Явление эпистаза генов
Необходимо ответить тот факт, что очень редко развитие того или иного признака определяется одним геном. В большинстве случаев какой – либо ген в единственном экземпляре становится фундаментом для развития различных признаков. Такое явление называется плейотропным взаимодействием генов. Также весьма распространено такое явление, когда один признак контролируется несколькими генами.
Определение 2
Неаллельные гены – это гены, которые расположены в различных местах (участках хромосом) и кодируют разные белки.
Статья: Эпистаз генов
Найди решение своей задачи среди 1 000 000 ответов
Неаллельные гены могут взаимодействовать между собой, когда один признак проявляется под действием нескольких генов.
В науке выделяется три формы взаимодействия генов между собой:
- комплементарность (или такое взаимодействие генов, которые дополняют друг друга и обуславливают развитие нового признака);
- полимерия или процесс генетического взаимодействия, при котором неаллельные гены влияют в одном напралвении на развитие одинакового признака;
- эпистаз (подавление гена иным геном).
Все указанные типы неаллельных взаимодействий встречаются с равной степенью вероятности и должны оцениваться с позиций гибридологического подхода.
ссылки
- Брукер Р.Дж. (2017). Генетика: анализ и принципы. McGraw-Hill Higher Education, Нью-Йорк, Нью-Йорк, США.
- Goodenough, U. W. (1984) Genetics. W. B. Saunders Co. Ltd, Пкиладелия, Пенсильвания, США.
- Griffiths A.J.F., Wessler R., Carroll S.B., Doebley J. (2015). Введение в генетический анализ (11-е изд.). Нью-Йорк: У. Х. Фриман, Нью-Йорк, Нью-Йорк, США.
- Miko, I. (2008) Epistasis: взаимодействие генов и фенотипические эффекты. Природоведческое образование 1: 197. nature.com
- Уайт Д., Рабаго-Смит М. (2011). Генотип-фенотипические ассоциации и цвет глаз человека. Журнал генетики человека, 56: 5-7.
- .
Что такое эпистаз?
Эпистаз – это феномен в генетике, который описывает вклад и взаимосвязь двух или более генных локусов в определении одного фенотипа. Другими словами, эпистаз можно определить как взаимодействие генов, при котором влияние одного аллеля гена влияет на действие аллелей другого гена. Например, если пигмент образуется под действием двух генов; ген 1 и ген 2, без экспрессии обоих генов, пигмент не может быть синтезирован, потому что ген 1 отвечает за производство промежуточной молекулы из молекулы-предшественника, а промежуточное соединение будет преобразовываться в пигмент посредством экспрессии гена 2. Следовательно, взаимосвязь между двумя генами требуется для окончательного производства пигмента, который дает фенотип. Это известно как эпистаз. Эпистаз также может использоваться для обозначения генов, которые маскируют эффекты другого гена.
Мутация одного гена или две мутации в локусах гена могут по-разному влиять на фенотип. Эпистаз можно разделить на различные формы, такие как положительный эпистаз, отрицательный эпистаз, антагонистический эпистаз и синергический эпистаз, в зависимости от мутаций и величины.
Примеры эпистаза
Так, рецессивные гены окраски собак из локуса С не позволяют синтезироваться пигментам, определяющим окраску шерсти.
Собака гомозиготная по ним — белая.
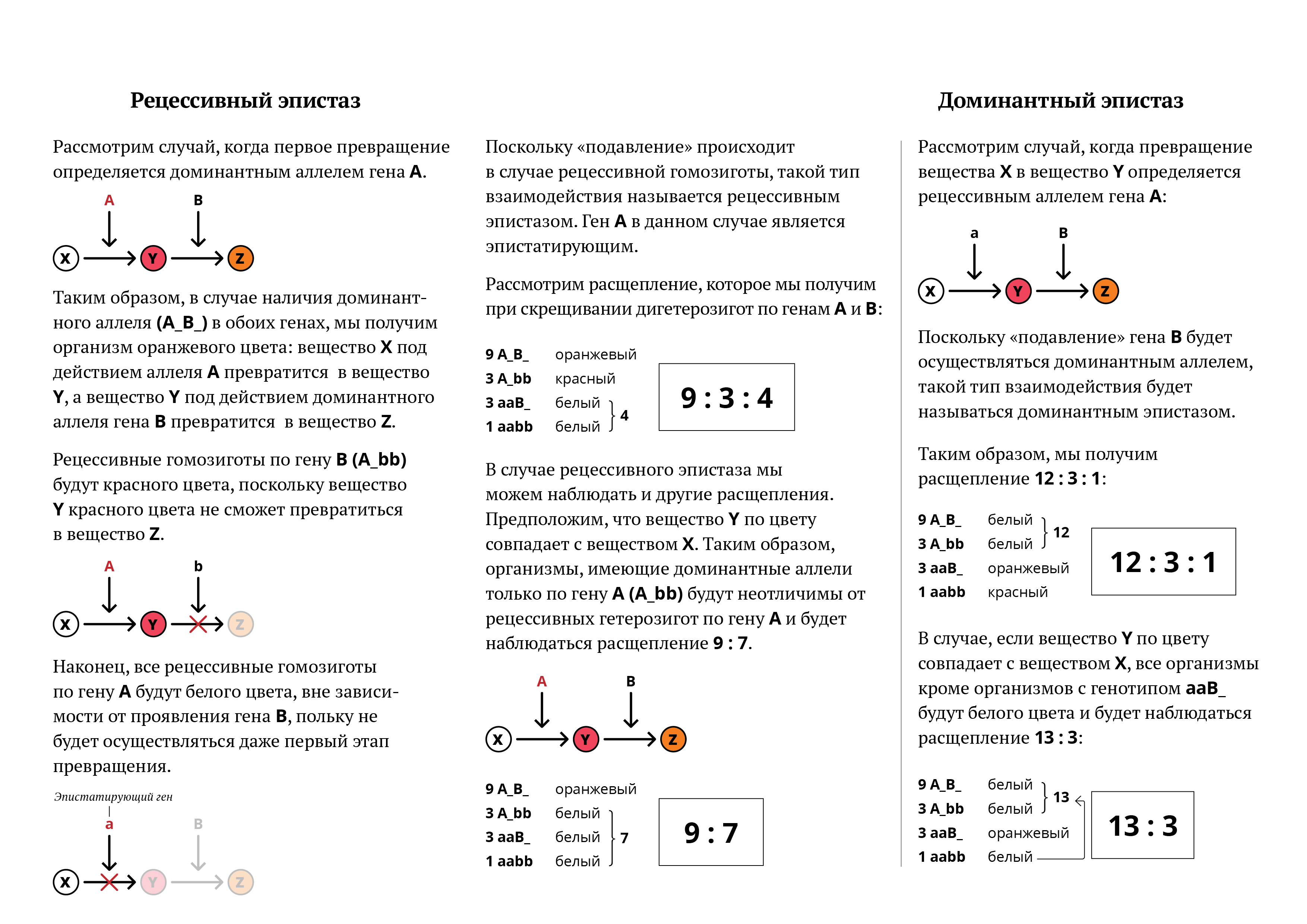
Такой тип эпистаза называется рецессивным. Скрещивание белой черноносой собаки с коричневой приведет к рождению в первом поколении гетерозиготных черных собак, а скрещивание гибридов F, между собой — к рождению 9 черных, 3 коричневых и 4 белых, т. е. будет наблюдаться отклонение от нормального расщепления 9:3:3:1.
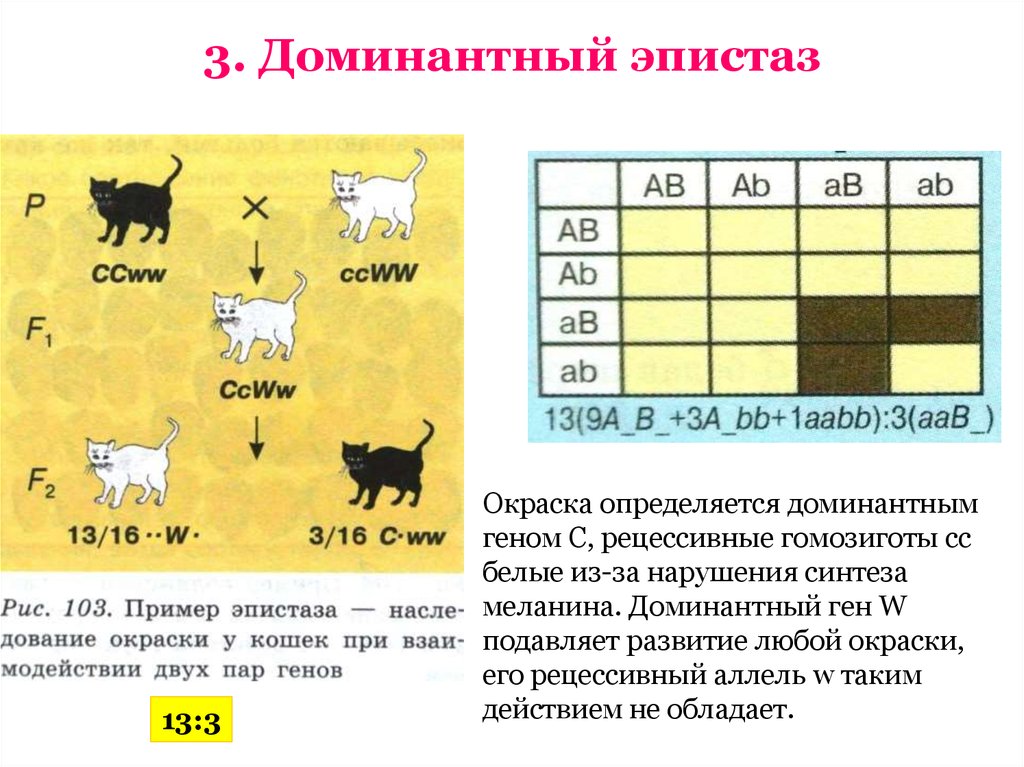
Скрещивание белой кошки, имеющей доминантный белый окрас, обусловленный доминантным аллелем W, с черной, напротив, приведет к рождению в первом поколении белых гетерозиготных котят.
Расщепление в F2 даст вместо ожидаемых 9:3:3:1 соотношение 12:3:1, где 12 — белых, 3 — полосатых и 1 черный котенок. Такой тип эпистаза называется доминантным. Отсутствие необходимых данных о роли первичных продуктов многих генов (ферментов, пептидов) в формировании сложных признаков, часто не позволяет точно установить характер взаимодействия неаллельных локусов, участвующих в биохимических процессах и составляющих основу образования этих признаков.
В одних случаях развитие признака при наличии двух неаллельных генов в доминантном состоянии рассматривают как комплементарное воздействие, в других — неразвитие признака, определяемого одним из генов при отсутствии другого гена в доминантом состоянии, расценивают как рецессивный эпистаз; если же признак развивается при отсутствии доминантного аллеля неаллельного гена, а в его присутствии не развивается, говорят о доминантом эпистазе.
На модификацию дигибридного скрещивания при рецессивном эпистазе
В брак вступают супруги, имеющие кровь II группы, матери которых имели I группу крови, и гетерозиготные по гену рецессивного эпистаза. Определите вероятные группы крови детей с учетом рецессивного эпистаза?
Решение
Обозначим гены:
| Ген | Признак |
| I0- 1А- 1В- 1А1В- хх>1А- хх>1в- хх>1А1в- X — х — | Iгруппа крови II группа крови III группа крови IV группа крови I гр. кр. I гр. кр. Рецессивный эпистаз I гр. кр. нормальный ген рецессивный эпистатический ген. |
Определите вероятные группы крови детей с учетом рецессивного эпистаза?
Фенотипы родителей:
II гр. крови — II гр. крови
гетерозигот. по 2 генам гетерозигот. по 2 генам
Ответ:_Расщепление по фенотипу 9:7, вероятность рождения со второй группой крови составляет 9/16, с первой 7/16, в 3/16 (IAIAcx, IAI°xx, IAI°xx) проявилась I группа крови вследствие рецессивного эпистаза и в 4/16 за счет сочетания гамет в зиготе (см. решетку Пеннета).
Механизмы эпистаза генов
Гены – супрессоры обладают способностью подавлять действие других генов, гены ингибиторы оказывают аналогичное действие. Реакция, возникающая от такого взаимодействия, может проявиться в рецессивном и доминантном вариантах.
Доминантный эпистаз наблюдается в том случае, если доминантная аллель гена – супрессора подавляет действие другого гена. Такая аллель, как правило, обозначается буквой I.
В свою очередь рецессивный эпистаз наблюдается в диаметрально противоположном случае при угнетении одного гена другим.
Фенотипическое расщепление при каждом виде эпистаза существенно различается. При доминантном эпистазе в случае скрещивания дигетерозигот расщепление может быть в следующих соотношениях:
- тринадцать к трем и семь к шести к трем;
- двенадцать к трем и к одному.
При рецессивном эпистазе расщепление может быть в следующих соотношениях:
- девять к трем и к четырём;
- тринадцать к трем.
Взаимодействие по типу эпистаза противоположно комплементарному. При наличии комплиментарного взаимодействия наблюдается дополнение одного гена другим. Эпистаз очень схож с доминированием, но разница проявляется в участие аллеля, принадлежащего к аллеморфной паре.

![Глава iii. наследование при моногенных различиях между исходными формами [1991 орлова н.н. - генетический анализ]](https://otlichaet.com/wp-content/uploads/4/9/b/49b4f9cc1ad8f0a43056490b6e1fa951.jpeg)




При эпистазе аллель одного гена подавляет проявление аллеля из другой аллеломорфной пары, т. е. неаллельного гена. Фенотипически эпистаз выражается в отклонении от расщепления, которое ожидается от стандартного наследования. Следует отметить тот факт, что нарушения законов Менделя в данном случае не происходит, поскольку распределения аллелей всех взаимодействующих генов соответствует закону независимого комбинирования тех или иных признаков.
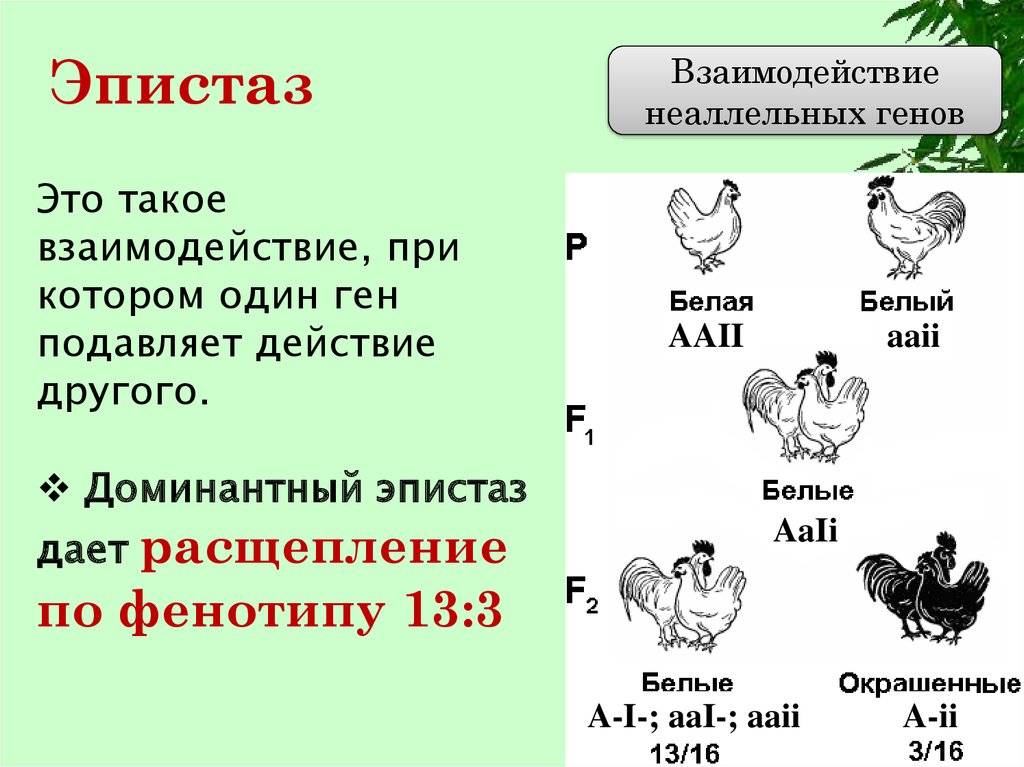
Эпистатическая система наследования обнаружена у кур. Некоторые породы кур имеют белое оперение (плимутрок, леггорн), другие породы имеют окрашенное оперение (полосатый плимутрок). При этом белое оперение различных пород кур определяется несколькими различными генами. Например, белая доминантная окраска определяется генами CCII, а рецессивная белая – ccii. Возможно и другое условное обозначение.
При этом ген С позволяет определить наличие предшественника пигмента или исследовать окрашенность пера и гены, отвечающие за не окрашенность пера. При скрещивании между собой гибридов F1 во втором поколении имеет место расщепление по окраске в отношении 13 белых: 3 окрашенных.
Также можно привести и другие примеры эпистатического взаимодействия генов. У человека существует так называемый бомбейский феномен. Люди, которые гетерозиготные по гену FUTI обладают нарушенной экспрессией генов, которые кодируют ферменты, отвечающие за образование эритроцитов A и B. На эритроцитах вместо нормальных генов образуются дефектные гены. Эти люди имеют группу крови O независимо от аллелей генов, которые определяют тип группы крови по системе ABO, не имеют антиегна A, антигена B.
Гены, которые определяют окраску шерсти у мышей также обладают эпистатическим взаимодействием. Окраска агаути у мышей обусловлена чередованием кольцевых пигментных полос по длине каждой части волосяного покрова. Она доминирует над чёрной окраской и определяется рецессивным мутантным аллелем. Если рецессивная мутация имеется в другом локусе, то пигментация полностью исчезает, и эта ситуация совершенно не зависит от того, какими аллелями представлен ген A.
Доминантный эпистаз у тыквы проявляется в окраске плодов. Доминантный аллель тыквы обуславливает ее белую окраску, вне зависимости от того, какими аллелями представлен ген B. Если локус A представлен рецессивной аллелью, то растения с генотипами BB и Bb имеют жёлтые плоды, а bb — зелёные.
Встречаются ситуации, когда наблюдается двойной эпистаз. Он встречается в тех случаях, когда встречаются гены, кодирующие ферменты одного метаболического пути. Если внутри клетки не будет создаваться нормальная форма одного из ферментов, то также не будет образовываться промежуточный продукт, который является фундаментом для всех последующих стадий. В результате нормативного продукта не будет образовываться при любой комбинации аллелей.
Также в качестве примера можно привести гены мухи – дрозофилы, которые кодируют ферменты – катализаторы, участвующие в превращении триптофана, который приобретает бурую окраску во время химических превращений. Если в процессе будет участвовать только рецессивный ген, то фенотип мух по окраске будет существенно отличаться от дикого типа.
Таким образом, эпистатическое взаимодействие неаллельных генов является одним из примеров подавления одного гена другим. Он позволяет отследить закономерности наследования генов и отразить специфику их взаимодействия внутри генотипа.
Эпистатическое действие генов
Окраска агути (рыжевато-серая) определяется доминантным геном А, его рецессивный аллель а в гомозиготном состоянии определяет черную окраску. Доминантный ген другой нары С определяет развитие пигмента, а гомозиготы по его рецессивному аллелю с являются альбиносами (отсутствие пигмента в шерсти и радужной оболочке глаз).
Особи, являющиеся доминантными гомозиготами по гену С или гетерозиготные по этому гену, будут иметь окраску шерсти в соответствии со своим генотипом по гену А (например, ААСС и ААСс будут рыжевато-серые агути, ааСС и ааСс — черные). Но все особи, гомозиготные по рецессивному гену-супреесору (ААсс, Аасс и аасс), будут альбиносами.
Как известно, доминирование есть подавление действия одной аллели другой аллелью, представляющих собой один ген: А > а, В > b, С > с и т. д.
Но существует взаимодействие, при котором аллель одного из генов подавляет действие аллелей других генов, например А > В или В А, а > В или b > А и т. д. Такое явление «доминирования» между генами называется эпистазом.
Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному взаимодействию.
Гены, подавляющие действие других генов, называются супрессорами, или ингибиторами.
Они могут быть как доминантными, так и рецессивными. Гены-супрессоры известны у животных (млекопитающие, птицы, насекомые) и у растений. Обычно они обозначаются I или Su в случае доминантного состояния генов и i или su для их рецессивных аллелей (от английских слов inhibitor или supressor).
В настоящее время эпистаз делят на два типа: доминантный и рецессивный.
Под доминантным эпистазом понимают подавление доминантной аллелью одного гена действия аллельной пары другого гена.
Из многих примеров доминантного эпистаза, установленных как на животных, так и на растениях, приведем лишь отдельные.
Некоторые породы кур имеют белое оперение (белый леггорн, белый примутрок и др.), другие же породы имеют окрашенное оперение (австралорп, ньюгемпшир, полосатый плимутрок и др.). Белое оперение разных пород кур определяется несколькими различными генами.
Так, например, доминантная белая окраска определяется генами CCII (белые леггорны), а рецессивная белая — ccii (белые суссексы, белые минорки, белые плимутроки). Ген С определяет наличие предшественник пигменту (хромогена), т. е. окрашенность пера, его аллель с — отсутствие хромогена и, следовательно, неокрашенность пера птицы. Ген I является подавителем действия гена С, аллель i не подавляет его действия. В присутствии даже одной дозы гена I в генотипе птицы действие генов окраски не проявится.
Поэтому при скрещивании белых леггорнов CCII с цветными породами CCii, как правило, доминирует белая окраска CCIi. При скрещивании белых плимутроков ccii с окрашенными породами CCii гибриды оказываются окрашенными Ccii. Следовательно, у леггорнов белая окраска является доминантной, а у плимутроков — рецессивной.
Если же производится скрещивание белых леггорнов CCII с рецессивными белыми плимутроками ccii, то в первом поколении цыплята оказываются тоже белыми CcIi.
При скрещивании между Собой гибридов F1 во втором поколении имеет место расщепление по окраске в отношении 13/16 белых : 3/16 окрашенных.
Наследование окраски у кур при взаимодействии двух пар генов (эпистаз)
Как можно объяснить полученное отношение?
Прежде всего, данное отношение говорит о расщеплении по двум генам, это расщепление можно представить как 9(С—I—) + 3(ccI—) + 1(ccii) = 13 и 3(С—ii), что соответствует формуле 9:3:3:1.
Очевидно, в этом случае окраска леггорнов обязана не присутствию особых генов белой окраски, а действию гена — подавителя окраски (I—). Тогда генотип гомозиготных белых леггорнов должен быть ССII, где I является геном — подавителем окраски, а С — геном окраски.
Предпосылки
Концепция доминирования была введена Грегором Иоганном Менделем. Хотя Мендель, «отец генетики», впервые использовал этот термин в 1860-х годах, он не был широко известен до начала двадцатого века. Мендель заметил, что для множества признаков садового гороха, связанных с внешним видом семян, семенных коробочек и растений, существует два дискретных фенотипа, такие как круглые или морщинистые семена, желтые или зеленые семена, красные и белые цветы или высокие растения по сравнению с низкорослыми. При раздельном разведении растения всегда давали одни и те же фенотипы, поколение за поколением. Однако когда линии с разными фенотипами скрещивались (скрещивались), у потомства обнаруживался один и только один родительский фенотип (зеленый, круглый, красный или высокий). Однако, когда эти гибридные растения были скрещены, растения-потомки показали два исходных фенотипа в характерном соотношении 3: 1, причем наиболее распространенным фенотипом был фенотип родительских гибридных растений. Мендель рассуждал, что каждый родитель в первом скрещивании был гомозиготным по разным аллелям (один родительский AA и другой родительский aa), что каждый вносил один аллель в потомство, в результате чего все эти гибриды были гетерозиготами (Aa), и что один из двух аллелей в гибридном кроссе доминировал над экспрессией другого: A замаскированный a. Окончательное скрещивание двух гетерозигот (Aa X Aa) даст потомство AA, Aa и aa в соотношении генотипов 1: 2: 1, причем первые два класса демонстрируют фенотип (A), а последний – фенотип (a)., таким образом, обеспечивая соотношение фенотипов 3: 1.
Мендель не использовал термины ген, аллель, фенотип, генотип, гомозигота и гетерозигота, все из которых были введены позже. Он ввел обозначения прописных и строчных букв для доминантных и рецессивных аллелей, соответственно, которые используются до сих пор.
В 1928 году британский популяционный генетик Рональд Фишер предположил, что доминирование действует на основе естественного отбора за счет вклада генов-модификаторов. В 1929 году американский генетик Сьюэлл Райт ответил, заявив, что доминирование – это просто физиологическое следствие метаболических путей и относительной необходимости задействованного гена. Объяснение Райта стало установленным фактом в генетике, и дебаты в основном прекратились. Однако на некоторые черты могут влиять эволюционные механизмы.
Хромосомы, гены и аллели
Большинство животных и некоторые растения имеют парные хромосомы и описываются как диплоид. У них есть две версии каждой хромосомы, одна из которых внесена яйцеклеткой матери, а другая – сперматозоидом отца, известная как гаметы, описанная как гаплоидная, и создан посредством мейоза. Затем эти гаметы сливаются во время оплодотворения во время полового размножения в новую одноклеточную зиготу, которая многократно делится, в результате чего возникает новый организм с таким же количеством пары хромосом в каждой (негаметной) клетке в качестве ее родителей.
Каждая хромосома из совпадающей (гомологичной) пары структурно подобна другой и имеет очень похожую последовательность ДНК (локусов, сингулярный локус). ДНК в каждой хромосоме функционирует как серия дискретных генов, которые влияют на различные признаки. Таким образом, каждый ген также имеет соответствующий гомолог, который может существовать в разных версиях, называемых аллелями. Аллели одного и того же локуса на двух гомологичных хромосомах могут быть идентичными или разными.
группа крови человека определяется геном, который создает группу крови A, B, AB или O и расположен в длинном плече хромосомы. девять. Есть три разных аллеля, которые могут присутствовать в этом локусе, но только два могут присутствовать у любого человека: один унаследован от их матери, а другой – от отца.
Если два аллеля данного гена идентичны, организм называется гомозиготным и считается гомозиготным по этому гену; если вместо этого два аллеля различны, организм является гетерозиготным и гетерозиготным. Генетический состав организма, находящийся либо в одном локусе, либо по всем его генам вместе, называется его генотипом. Генотип организма прямо или косвенно влияет на его молекулярные, физические и другие характеристики, которые по отдельности или вместе называются его фенотипом. В гетерозиготных генных локусах эти два аллеля взаимодействуют, создавая фенотип.
Генотип как целостная система. Взаимодействие генов
Вы познакомились с основными закономерностями наследования признаков у организмов.
Знания о цитологических основах наследственности развивались постепенно. Так, Г.Мендель, ничего не зная о генах и хромосомах, гениально сформулировал следующие положения:






- каждый признак организма контролируется парой наследственных задатков, или факторов (сейчас их называют генами);
- наследственные факторы (гены) могут существовать в двух состояниях: доминантном и рецессивном;
- гаметы генетически чисты, то есть содержат по одному наследственному фактору (гену) из каждой аллельной пары;
- при образовании зиготы наследственные факторы (гены) не смешиваются, остаются в «чистом» виде.
В 1909 г. Иогансен заменил термин «фактор» термином «ген».
В 1910-1920 гг. американский ученый Томас Морган сформулировал хромосомную теорию наследственности, согласно которой гены располагаются в хромосомах в линейном порядке, каждый ген занимает в хромосоме определенное место и влияет на формирование определенного признака.
Дальнейшие исследования внесли существенные поправки и дополнения в хромосомную теорию наследственности и углубили знания о гене.
Большинство генов влияет не на один признак организма, а на несколько, то есть обладает множественным действием.
Например, один из рецессивных генов у мухи дрозофилы обусловливает белую окраску глаз, низкую плодовитость и небольшую продолжительность жизни. У мышей под влиянием рецессивного гена в потомстве появляются карликовые особи, которые отличаются от нормальных мышей более тупыми мордочками, короткими усиками и хвостами, меньшей продолжительностью жизни.
В генотипе многих организмов имеются рецессивные летальные гены, одним из проявлений которых является гибель организма до полного завершения его развития, если летальный ген находится в гомозиготном состоянии.
Например, ген, обусловливающий отсутствие хлорофилла, приводит к гибели гомозиготных рецессивных проростков кукурузы.
В настоящее время установлено, что гены в генотипе тесно взаимосвязаны и взаимодействуют. Поэтому генотип рассматривают не как арифметическую сумму генов, а как целостную систему взаимодействующих генов. Полное и неполное доминирование обусловлено взаимодействием двух аллелей одного гена (А и а или B и b), которые определяют проявление у организма какого-то одного признака.
Это аллель 1000 ное взаимодействие генов.
Существуют и другие разнообразные взаимодействия генов. Так, гены-модификаторы ослабляют или усиливают действие других генов.
От их взаимодействия зависит, например, пятнистая окраска шерсти у животных: у одних особей пятнистость выражена в большей степени, у других в меньшей.
Некоторые неаллельные гены, одновременно находясь в генотипе, обусловливают развитие нового признака у организма. Например, при скрещивании черного и белого кроликов появляется потомство, которое имеет серую окраску шерсти.
При скрещивании гибридных особей между собой в потомстве происходит необычное расщепление по фенотипу: 9 серых : 3 черных : 4 белых.
Почему при скрещивании черного и белого кроликов в потомстве появляется новый признак — серая окраска шерсти?
Дело в том, что у кроликов за окраску шерсти отвечает аллельная пара генов Аа (А — черная, а — белая окраска шерсти), а за распределение пигмента по длине волоса — аллельная пара генов Вb (В — пигмент находится у корня волос, b — пигмент распределяется равномерно по длине волоса).
Теперь понятно, что черными будут кролики, у которых в генотипе ген А сочетается с геном b (Aabb, Aabb).
Кролики с генотипами ааВb, aaBB, aabb будут белыми, так как пигмент у них отсутствует. Серая окраска шерсти появляется только в том случае, если в генотипе соединились два доминантных неаллельных гена — А и В.
Это значит, что в клетках образуется пигмент (есть ген А), но он скапливается у основания волос (АаВb, ААВb, AaBB, AABB).
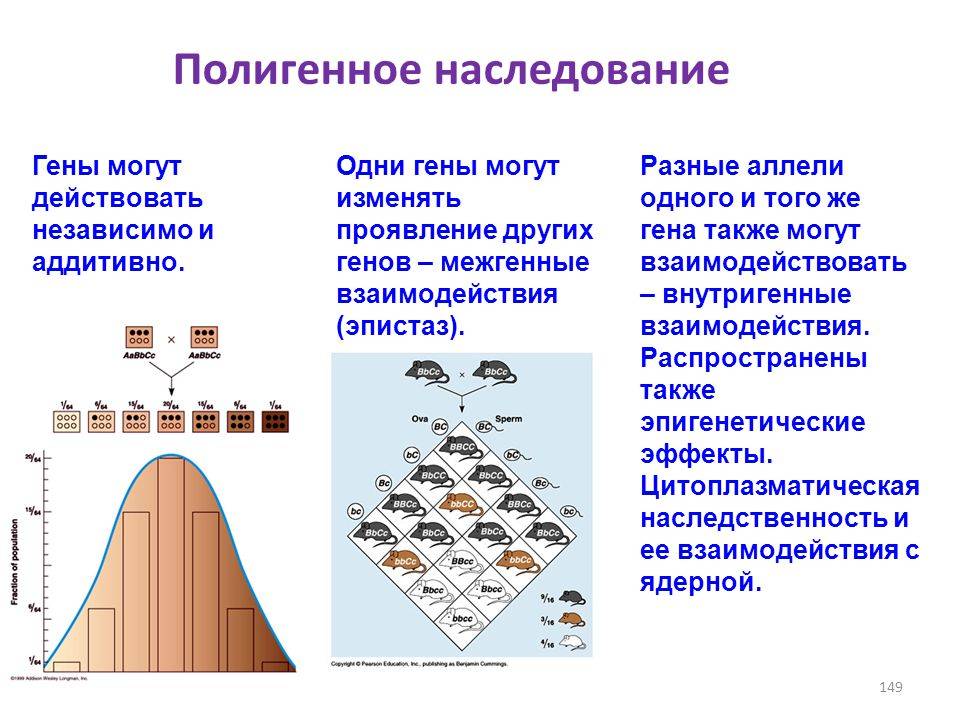
Большинство признаков организма формируется благодаря взаимодействию нескольких генов, которые отвечают за один и тот же признак, но не являются аллельными.
К таким признакам относят рост, массу тела животных, жирность и надои молока коров, яйценоскость кур. Эти гены принято обозначать одной и той же буквой со значками 1, 2, 3 и т. д. Цвет кожи у человека, например, обусловлен двумя неаллельными генами A1 и А2. Он зависит от числа доминантных генов в генотипе: чем их больше, тем темнее кожа. Люди, генотип которых А1A1А2А2, будут иметь темную кожу, а кожа людей с генотипом а1а1а2а2 светлая.
Люди с генотипами А1a1А2a2, А1a1a2a2 имеют различные оттенки кожи — от более темной до почти светлой.
